Special Issue: Molecular System BioenergeticsThis special issue belongs to the section Biochemistry, Molecular Biology and BiophysicsInternational Journal of Molecular Sciences (ISSN 1422-0067) |
|---|
[Editors] [Feature Papers] [Announced Papers] [Published Papers] [List of Keywords] [Manuscript Submission]
Manuscript
Submisison Deadline: 31 January 2009
Editors:
Editorial Office
IJMS Editorial Office
Molecular Diversity Preservation International
Kandererstrasse 25, CH-4057 Basel, Switzerland
Tel. +41-61-683-7734; Fax: +41-61-302-8918
Address manuscript submissions to: Dr. Yuan Gao, E-mail: [email protected]
Guest Editor
Prof. Dr. Valdur A. Saks
Laboratory of Bioenergetics, Joseph Fourier University, 2280, Rue de la Piscine, BP53X-38041, Grenoble Cedex 9, France
Tel. 00-33-4-7663-5627; Fax: 00-33-4-7651-4218
E-mail: [email protected]
Rationale for the special issue on "Molecular System Bioenergetics"
Dear Colleagues,
Valdur A. Saks
IJMS Editorial Office
Molecular Diversity Preservation International
Kandererstrasse 25, CH-4057 Basel, Switzerland
Tel. +41-61-683-7734; Fax: +41-61-302-8918
Address manuscript submissions to: Dr. Yuan Gao, E-mail: [email protected]
Guest Editor
Prof. Dr. Valdur A. Saks
Laboratory of Bioenergetics, Joseph Fourier University, 2280, Rue de la Piscine, BP53X-38041, Grenoble Cedex 9, France
Tel. 00-33-4-7663-5627; Fax: 00-33-4-7651-4218
E-mail: [email protected]
Rationale for the special issue on "Molecular System Bioenergetics"
Dear Colleagues,
This Special Issue continues the series of publications on application
of the new strategy of research – Systems Biology- in an important area
of biological research: for investigation of the mechanisms of
regulation of integrated processes of energy metabolism of cells. This
series was started by publication by Wiley VCH, Weinheim, Germany in
2007 of the book “Molecular System Bioenergetics. Energy for Life” (http://www3.interscience.wiley.com/cgi-bin/bookhome/117349267).
Systems Biology is a new paradigm of biological sciences which opens
wide perspectives of better understanding of complex biological
processes at different levels, introducing network theories and new
concepts such as that of system level properties not predictable from
the studies of isolated components of the cells. Examples of these
important system level properties are the phenomena of metabolic
compartmentation and functional coupling depending on specific
intracellular organization. This concept is also central for
understanding the mechanisms of regulation of cellular energetics and
other metabolic processes in the cells in vivo.
In the current Specific Issue the authors describe and analyze wide
variety of different aspects of the current state of the art in this
important area, starting with description of philosophical and
historical basis of systems biology approaches, network theories and
their applications in biology and in particular in bioenergetics,
compartmentation phenomena, intracellular interactions and mechanisms
of signalling, important role of the cytoskeleton, in particular in the
control of mitochondrial dynamics, arrangement, function and in modular
organization of energy metabolism.
Valdur A. Saks
Keywords:
- systems biology
- molecular and cellular bioenergetics
- cytoskeleton
- compartmentation
- integrated energy metabolism
- intracellular signalling
Manuscript Submission:
You are invited to submit your manuscript by e-mail to [email protected]
with copy to the guest editor. Papers will be published continuously
until the deadline and listed together at this special issue website.
Manuscript Submisison Deadline: 31 January 2009
Manuscript Submisison Deadline: 31 January 2009
The International Journal of Molecular Sciences (http://www.mdpi.org/ijms/;
ISSN 1422-0067) was launched in 2000 and is an Open Access journal.
According to SCI database, the IJMS has an impact factor (cites per
article) of 0.750 in 2007. The IJMS maintains a rapid editorial
procedure and a rigorous peer-review system. Well written papers have
been peer-reviewed and published in less than 4 weeks from manuscript
submission. This journal is abstracted and indexed very rapidly by
Chemical Abstracts, the Science Citation Index Expanded, the Chemistry
Citation Index, EMBASE, Scopus and Google Scholar. The IJMS has been
accepted for coverage in PubMed/MEDLINE starting the 2008 volume.
Articles from this volume will appear in PubMed by end of November
2008. Additionally, all articles will be archived in PubMed Central.
Submitted papers should not have been published previously, nor be currently under consideration for publication elsewhere. Please visit the Instructions for Authors at http://www.mdpi.org/ijms/publguid.htm before submitting a paper. Open Access publication fees are 800 CHF per paper. English correction fees (250 CHF) will be added in certain cases (1050 CHF per paper for those papers that require extensive additional formatting and/or English corrections).
Submitted papers should not have been published previously, nor be currently under consideration for publication elsewhere. Please visit the Instructions for Authors at http://www.mdpi.org/ijms/publguid.htm before submitting a paper. Open Access publication fees are 800 CHF per paper. English correction fees (250 CHF) will be added in certain cases (1050 CHF per paper for those papers that require extensive additional formatting and/or English corrections).
Feature Papers:
Announced Papers:
Published Papers:
Author: Denis Noble
Affiliation: CBE FRS, Balliol College, OX1 2BJ, and Department of Physiology, Anatomy and Genetics, Parks Road, Oxford OX1 3PT, UK. E-mail: [email protected]; Tel. +44-1865 272528; Fax +44-1865 272554. http://musicoflife.co.uk/
Title: (title to come)
Authors: Valdur Saks and Claire Monge
Affiliation: INSERM U884, Laboratoire de la Bioénergétique et Appliquée, Université Joseph Fourier, 2280 Rue de la Piscine, BP 53, Grenoble Cedex 9, France. E-mail : [email protected]; Tel. +33-476635627; Fax : +33-476514218.
Type: Review
Title: Philosophical and Historical Basis of Systems Biology Approaches: from Hegel to Noble. Application for Bioenergetic Reasearch
Author: A. Kuznetsov
Affiliation: D. Swarovski Research Laboratory, Department of Transplant Surgery, Innsbruck Medical University (IMU), Innrain 66, A- 6020 Innsbruck, Austria. E-mail: [email protected]; Tel. +43-512504-27811; Fax: +43-512504-24625
Type: Review
Title: Heterogeneity of Mitochondria and Mitochondrial Function within Cells: Another Level of Mitochondrial Complexity
Authors: Aon Miguel and Sonia Cortassa
Affiliation: Johns Hopkins Univ. School of Medicine, Institute of Molecular Cardiobiology, 720 Rutland Ave. 1055 Ross Bldg., Baltimore, MD 21205, USA. E-mails: [email protected], [email protected]u; Tel. +1-(410) 955-2759; Fax: +1-(410) 955-7953
Type: Review
Title: Control and Regulation in Metabolic and Transport Networks of Integrated Cardiomyocycte Function
Authors: Dzeja Petras and André Terzic
Affiliation: Division of Cardiovascular Diseases, Departments of Medicine, Molecular Pharmacology and Expermental Therapeutics, Mayo Clinic College of Medicine, Guggenheim 7, Rochester, MN, 55905 USA. E-mail : [email protected], [email protected]; Tel. +1-507 2660606
Type: Article
Title: (title to come)
Authors: Marko Vendelin and J. Engelbrecht
Affiliation: Laboratory of Systems Biology, Institute of Cybernetics, Akadeemia tee 21, 12618 Tallinn, Estonia.E-mail: [email protected], [email protected]; Tel. 372 620 4160; Fax: 372 620 4151
Type: Article
Title: (title to come)
Authors: Jacques Demongeot
Affiliation: CBE FRS, Balliol College, OX1 2BJ, and Department of Physiology, Anatomy and Genetics, Parks Road, Oxford OX1 3PT, UK. E-mail: [email protected]; Tel. +44-1865 272528; Fax +44-1865 272554. http://musicoflife.co.uk/
Title: (title to come)
Authors: Valdur Saks and Claire Monge
Affiliation: INSERM U884, Laboratoire de la Bioénergétique et Appliquée, Université Joseph Fourier, 2280 Rue de la Piscine, BP 53, Grenoble Cedex 9, France. E-mail : [email protected]; Tel. +33-476635627; Fax : +33-476514218.
Type: Review
Title: Philosophical and Historical Basis of Systems Biology Approaches: from Hegel to Noble. Application for Bioenergetic Reasearch
Abstract:
In our days, we live in the time of change of the paradigm of
biological sciences. Reductionism that used to be the philosophical
basis of biochemistry and molecular biology during last six decades
when everything, from genes to proteins and organelles - were studied
in their isolated state is leaving its place to Systems Biology that
favours the study of integrated systems at all levels: cellular, organ,
organism, and population. Reductionism was justified in the initial
stages of biological research giving a wealth of information on system
components. It is timely topic to put them together and analyze them in
interaction, to understand the principles of functioning of the whole.
However, in historical perspective first systems biology approach was
applied already by Claude Bernard about 150 years ago (see ref. 1 for
review). These developments follow very precisely the dialectical
principles of development from thesis to antithesis to synthesis
discovered by Hegel (Scheme 1).
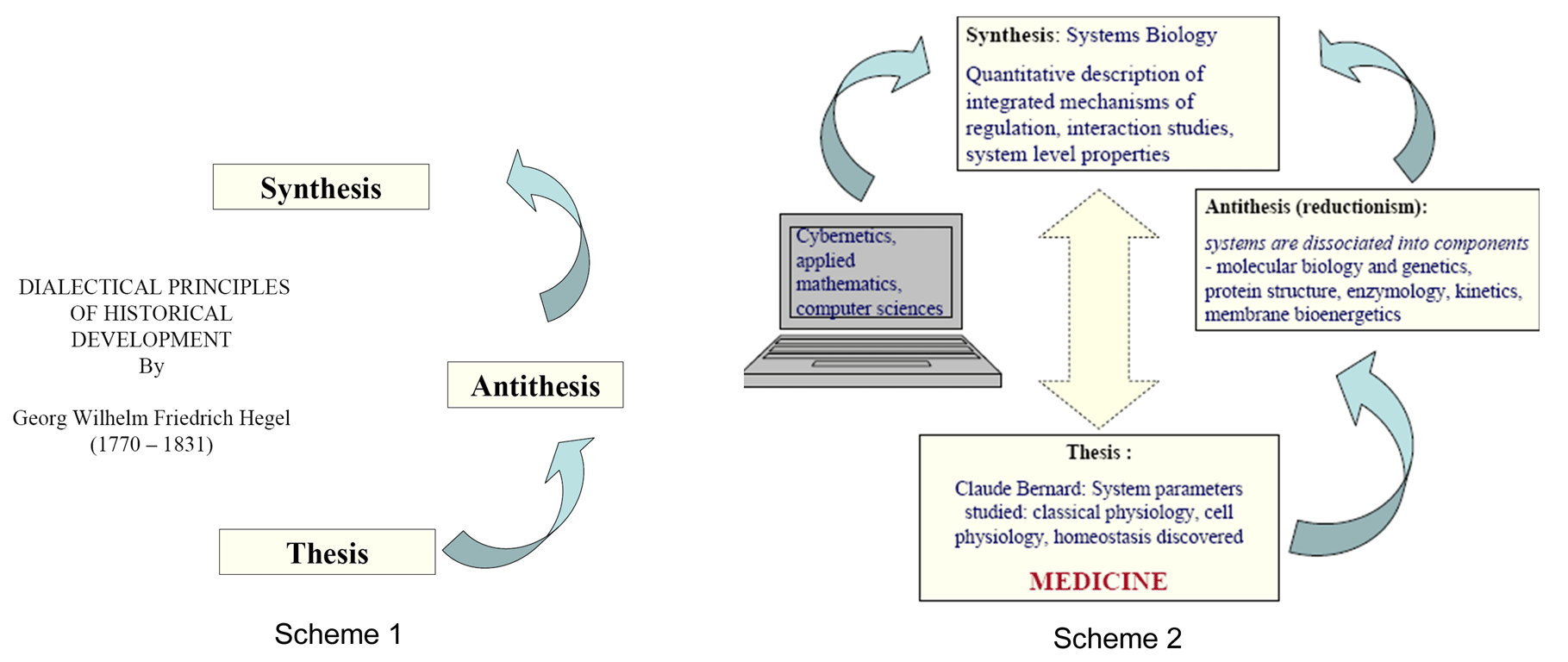
Dialectical principles of Hegel can be perfectly and very logically applied for description and analysis of the development of biological sciences (Scheme 2). The aim of Systems Biology is the higher – level analysis of complex biological systems (see ref. 2 and 3 for review) by using the wealth of information obtained in studies of isolated components, applying the methodological approaches of cybernetics, applied mathematics, network analysis, nonequilibrium thermodynamics of open systems (see ref. 4 for review) etc.
The Systems Biology opens new perspectives for studies of the integrated processes of energy metabolism in different cells (3). These integrated systems acquire new, system - level properties due to interaction of cellular components, such as metabolic compartmentation, channeling and functional coupling mechanisms, which are central for regulation of the energy fluxes. State of art of these studies in the new area of Molecular System Bioenergetics is analyzed.
References:
1. Noble D. 2008. Claude Bernard, the first systems biologist, and the future of physiology. Exp. Physiol. 93.1: 16 - 26
2. Kitano H. 2002. Systems Biology: A brief overview. Science, 295: 1662-1664.
3. Noble D. 2006. The music of life. Biology beyond the genome; Oxford University Press: Oxford, UK.
4. Prigogine I., Strengers I (1986) La Nouvelle Alliance. Les Editions Gallimard, Paris.
Dialectical principles of Hegel can be perfectly and very logically applied for description and analysis of the development of biological sciences (Scheme 2). The aim of Systems Biology is the higher – level analysis of complex biological systems (see ref. 2 and 3 for review) by using the wealth of information obtained in studies of isolated components, applying the methodological approaches of cybernetics, applied mathematics, network analysis, nonequilibrium thermodynamics of open systems (see ref. 4 for review) etc.
The Systems Biology opens new perspectives for studies of the integrated processes of energy metabolism in different cells (3). These integrated systems acquire new, system - level properties due to interaction of cellular components, such as metabolic compartmentation, channeling and functional coupling mechanisms, which are central for regulation of the energy fluxes. State of art of these studies in the new area of Molecular System Bioenergetics is analyzed.
References:
1. Noble D. 2008. Claude Bernard, the first systems biologist, and the future of physiology. Exp. Physiol. 93.1: 16 - 26
2. Kitano H. 2002. Systems Biology: A brief overview. Science, 295: 1662-1664.
3. Noble D. 2006. The music of life. Biology beyond the genome; Oxford University Press: Oxford, UK.
4. Prigogine I., Strengers I (1986) La Nouvelle Alliance. Les Editions Gallimard, Paris.
Author: A. Kuznetsov
Affiliation: D. Swarovski Research Laboratory, Department of Transplant Surgery, Innsbruck Medical University (IMU), Innrain 66, A- 6020 Innsbruck, Austria. E-mail: [email protected]; Tel. +43-512504-27811; Fax: +43-512504-24625
Type: Review
Title: Heterogeneity of Mitochondria and Mitochondrial Function within Cells: Another Level of Mitochondrial Complexity
Abstract:
Beyond fundamental role in energy metabolism mitochondria perform a
great variety of other important cellular functions. However, the
interplay among these roles of mitochondria is still poorly understood.
Importantly, mitochondria localized in different regions of a cell may
display different morphology (small spheres, short rod-like shape,
spaghetti-like shape, long branched tubules, complex mitochondrial
network), dissimilar biochemical properties, or may differently
interact with other intracellular structures. Recent advances in live
imaging technique have discovered also functional heterogeneity of
mitochondria in respect to mitochondrial redox state, membrane
potential, respiratory activity, uncoupling proteins, mitochondrial ROS
and calcium. Distinct mitochondrial subsets may also have different
responses to substrates and inhibitors and may vary in their
sensitivity to pathology, resistance to apoptosis, oxidative stress,
etc, demonstrating also heterogeneous dynamic motility, combining
different types of mitochondrial motion (e.g. small oscillatory
movements in mitochondrial network, filament extension, retraction,
fast and frequent oscillating branching and long-distance translocation
of single mitochondrion or mitochondrial filament). All these
observations show a new level of mitochondrial complexity and strongly
suggest that intracellular position and/or specific surrounding of
mitochondria within the cell may define their functional features,
suggesting also that different mitochondrial subpopulations, clusters
or even single mitochondrion may carry out diverse processes in a cell.
An important and still unresolved question is how heterogeneity of
mitochondrial function and regional specializations of mitochondria are
genetically defined and to which extent this heterogeneity may be
dependent on environmental aspects.
In addition, mitochondrial defects can be heterogeneously expressed in different mitochondrial subpopulations, which may be differently involved in pathological processes. They may have diverse sensitivities to injury or be associated with dissimilar metabolic consequences. The analysis of metabolic and functional diversity of mitochondria, thus, have important implications for analysis of mitochondria related pathologies like ischemia-reperfusion injury, oxidative stress, various cytopathies, etc. The application of mitochondrial imaging opens a very promising avenue for the development of new diagnostic approach for the detection of regional mitochondrial defects. Analysis of heterogeneity of mitochondria and mitochondrial function therefore represents a new challenging area in the mitochondrial research, potentially leading to the integration of mitochondrial bioenergetics and cell physiology with various physiological and pathophysiological implications.
In addition, mitochondrial defects can be heterogeneously expressed in different mitochondrial subpopulations, which may be differently involved in pathological processes. They may have diverse sensitivities to injury or be associated with dissimilar metabolic consequences. The analysis of metabolic and functional diversity of mitochondria, thus, have important implications for analysis of mitochondria related pathologies like ischemia-reperfusion injury, oxidative stress, various cytopathies, etc. The application of mitochondrial imaging opens a very promising avenue for the development of new diagnostic approach for the detection of regional mitochondrial defects. Analysis of heterogeneity of mitochondria and mitochondrial function therefore represents a new challenging area in the mitochondrial research, potentially leading to the integration of mitochondrial bioenergetics and cell physiology with various physiological and pathophysiological implications.
Authors: Aon Miguel and Sonia Cortassa
Affiliation: Johns Hopkins Univ. School of Medicine, Institute of Molecular Cardiobiology, 720 Rutland Ave. 1055 Ross Bldg., Baltimore, MD 21205, USA. E-mails: [email protected], [email protected]u; Tel. +1-(410) 955-2759; Fax: +1-(410) 955-7953
Type: Review
Title: Control and Regulation in Metabolic and Transport Networks of Integrated Cardiomyocycte Function
Authors: Dzeja Petras and André Terzic
Affiliation: Division of Cardiovascular Diseases, Departments of Medicine, Molecular Pharmacology and Expermental Therapeutics, Mayo Clinic College of Medicine, Guggenheim 7, Rochester, MN, 55905 USA. E-mail : [email protected], [email protected]; Tel. +1-507 2660606
Type: Article
Title: (title to come)
Authors: Marko Vendelin and J. Engelbrecht
Affiliation: Laboratory of Systems Biology, Institute of Cybernetics, Akadeemia tee 21, 12618 Tallinn, Estonia.E-mail: [email protected], [email protected]; Tel. 372 620 4160; Fax: 372 620 4151
Type: Article
Title: (title to come)
Authors: Jacques Demongeot
Affiliation:
Laboratoire TIMC-IMAG UMR 5525 - Pavillon Taillefer, Faculté de
Médecine de Grenoble - 38700 La Tronche. Tel. 33 (0)4 56 52
01 08; Fax: 33 (0)4 56 52 00 44
E-mail: [email protected]
Title: Robustness in Regulatory Networks. A Generic Approach with Applications at Different Levels: Physiologic, Metabolic and Genetic
E-mail: [email protected]
Title: Robustness in Regulatory Networks. A Generic Approach with Applications at Different Levels: Physiologic, Metabolic and Genetic
Abstract: The regulatory networks
are often studied on their dynamical side (existence of attractors, study of
their trajectory stability) but rarely on their robustness, that is their
ability to resist to external perturbations, then offering the same
spatio-temporal patterns independently of the loss of nodes or edges in their
interaction architecture, or of the change in their environmental boundary
conditions, or more of their mode of updating (e.g. co-expression versus
sequential expression in genetic networks). We will define precisely the generic
notion of critical node or edge in the interaction graph whose disappearance
can cause dramatic changes in the number and type of attractors (e.g. passage
from a bistable behavior to a unique periodic regime) or in the range of their
basins of stability. We will treat some examples: bulbar cardio-respiratory
physiologic regulation, cell cycle genetic control and feather morphogenesis
metabolic driving.
Announced Papers:
(none)
Published Papers:
(none)
© 2008 by MDPI, Basel, Switzerland